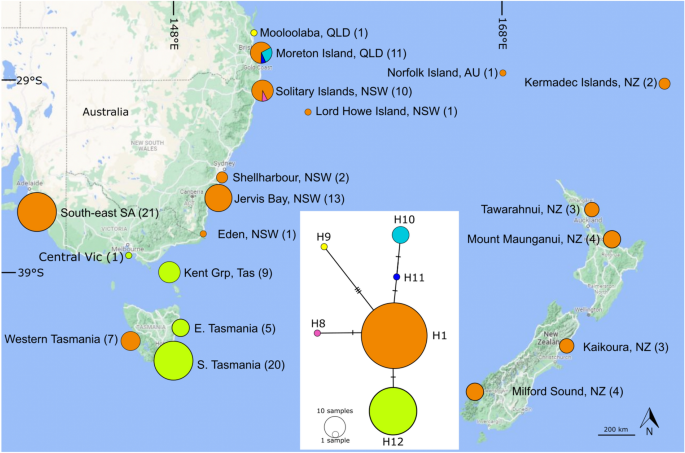
IntroductionClimate modification has actually driven modern decrease and loss of kelp forests worldwide (e.g. 1,2) with an accompanying loss of the significant environmental and financial worths that kelp forests provide3,4,5. Equatorward-range edge kelp populations are especially susceptible to environment modification as these areas are going through warming at or beyond kelp thermal tolerance thresholds6,7,8. Concerningly, these rear edge populations might consist of distinct adaptive9,10 or evolutionary6 hereditary variety that is likewise under danger. Kelp can in some cases continue at lower latitudes, assisted by cool water upwelling or in deep-water refugia where they are secured by thermoclines6,11,12,13,14. Recognition of these sanctuary locations is essential to make sure crucial hereditary variety is safeguarded and to comprehend how extant and previous environments shape types circulation and evolutionary variety. While modern hereditary variety can be constrained in low latitude, rear edge populations of temperate types (e.g. 5,8,14,15), evolutionary variety might be high due to historic processes16. Throughout the Pleistocene glaciation, lots of marine organisms were just able to continue within ice-free refugia at lower latitudes17,18,19. As temperature level and water level increased and glaciers pulled away, types quickly distributed poleward and recolonised brand-new locations. The signatures (‘ creator results’) of such distributional modifications can be seen in patterns of evolutionary hereditary variety with previous low latitude refugia frequently harbouring high hereditary variety. On the other hand, locations that were recently recolonised following termination or variety contractions show lower variety. Examples of low latitude glacial refugia with high hereditary variety can be discovered in temperate seaweeds consisting of Durvillaea antarctica (Chamisso) Hariot, 189220, Saccorhiza polyschides (Lightfoot) Batters, 190211 and Laminaria ochroleuca Bachelot de la Pylaie, 182421. Marine environments in Australia have a steady weather history and have not straight knowledgeable glaciation events22. Rather, Pleistocene cooling triggered water level to drop and isotherms to move equatorward around the Australian continent which have actually likewise left imprints on types evolutionary diversity23,24,25. A drop in sea level around southern Australia triggered the closure of Bass Straight resulting in seclusion in between the east and southern shorelines and hereditary signatures of refugia in lots of marine species26,27,28. The drop in water level and temperature level connected with glaciation would likewise have actually triggered vertical migration of types to much deeper depths as they followed thermal and light optima. Extensive terminations of rocky reef marine organisms were likewise most likely in locations with a narrow continental rack (e.g. the south eastern Australian shoreline) as sea levels fell and the accessibility of rocky reef significantly decreased (or ended up being non-existent) in lots of regions24. Ecklonia radiata (C. Agardh) J. Agardh, 1848 is the dominant and most extensively dispersed Laminarian kelp in the Southern Hemisphere2 and the only kelp throughout much of its variety, offering the dominant biogenic environment structure and supporting enormous environmental and financial values3. Previous research study on evolutionary variety in E. radiata in south-eastern Australia26 and Western Australia6 exposed hereditary variety to be low and controlled by a single cosmopolitan haplotype, which was likewise discovered in South Africa and Oman (where it is now extinct) 6, in addition to a couple of endemic local haplotypes. Durrant et al. 26 recommended that this low variety in south-eastern Australia was because of quick colonisation from the northern hemisphere instead of recolonisation following termination since no refugial locations with greater hereditary variety were discovered. Latitudes lower than 35 ° S on Australia’s east coast have the densest modern E. radiata forests29 and represents the location where Pleistocene temperature level would have been ideal for environment refugia to occur30 however have actually never ever been tested. The low latitudinal limitation of circulation of E. radiata on Australia’s east coast is at Moreton Island (27 ° S, Queensland) where this types just takes place on cooler, deep water reefs (30-80 m) listed below thermoclines14,31. This area is likewise fascinating from a historic environment modification viewpoint. Pleistocene glaciation triggered big drops in water level and temperature level that would likely have actually triggered substantial shifts in both the latitudinal and vertical circulation, consisting of colonisation of what are presently much deeper rocky reefs by E. radiata. At this time the Moreton Island area would have experienced ideal temperature levels (21– 24 ° C )to support E. radiata populations30. Prevalent regional terminations were likewise most likely as the currently narrow continental rack (and accessibility of rocky reef) ended up being considerably lowered south of Moreton Island with a fall in sea level24,32,33. The extant deep water Moreton Island populations might represent both a historic and modern haven from altering environments and harbour high hereditary variety. Ecklonia radiata likewise traditionally happened on the separated Lord Howe Island (31 ° S) with 2 herbarium specimens verifying its presence there. Even more north, a remote deep-water population of E. radiata exists east of Norfolk Island (29 ° S) 34 where a single herbarium sample exists (Auckland Museum– AK146440) from 1930, and a current specimen was dredged up throughout fishing (Mark Scott, pers. comm). There have actually been no modern verifications of extant populations on Lord Howe Island in spite of extensive environment studies. We presume that this population is extinct. This low latitude island might likewise represent a lost refugia that when harboured distinct or high hereditary variety. Considered that eastern Australia is warming at a rate that is around 4 times higher than the worldwide average35,36,37 which has actually driven modern decreases in E. radiata7, it concerns recognize evolutionary refugia to much better comprehend how environment drives the circulation of this vital types and to secure special hereditary variety. While present understanding of E. radiata population genes shows that these low latitude populations have low neutral hereditary variety due to minimal extant connectivity15,38 however have actually been chosen for greater thermal tolerance10, it stays unidentified how historic procedures have actually formed this structure types. We utilize the mitochondrial Cytochrome C Oxidase (COI) marker to analyze evolutionary hereditary (haplotype) variety amongst extant and putatively extinct E. radiata populations from throughout its complete latitudinal variety in eastern Australia and throughout the south-west Pacific. We anticipated that haplotype variety would be greater at lower latitudes due to their prospective function as historic refugia. Products and methodsEcklonia radiata specimensTo take a look at patterns of evolutionary variety of Ecklonia radiata throughout the south-west Pacific, we utilize a mix of newly gathered E. radiata (gathered as in Ref.15) and accessed herbarium specimens of unusual (Norfolk Island; just one sample in herbaria worldwide) and the putatively extinct population at Lord Howe Island (just 2 samples in herbaria worldwide), (Supplementary Information, see Fig. 1 for places). We acquired subsamples of dried herbarium specimens of E. radiata from Norfolk Island gathered in 1930 (Auckland Museum, New Zealand (NZ), accession AK146440), and Lord Howe Island, gathered in June 1933 (National Herbarium of New South Wales (NSW), Sydney, accession NSW817202). We likewise got a Lord Howe Island sample gathered in 1966 (Melbourne Herbarium MEL16690 however it did not produce DNA that enhanced. We likewise accessed thallus and stipe product from a single deep-water specimen, gathered from 70 m off Moreton Island, QLD in August 2017 (this is the only sample readily available in herbaria) that was held at the QLD herbarium (AQ1019163) repaired in 10% formalin, and 2 deep water, dried herbarium specimens from the Kermadec Islands (Auckland Museum, NZ). Figure 1Haplotype network and haplotype circulation of Ecklonia radiata based upon COI barcode markers from the south-western Pacific. Colours represent various haplotypes; circle size is agent of sample size (sample size likewise offered in parentheses after tasting place name) and network link hatch marks represent base set replacements. Haplotype numbers represent Coleman et al. 6. Map developed in Inkscape v1.2 (https://inkscape.org/release/inkscape-1.2/). Complete size image Fresh samples of Ecklonia radiata were gathered from numerous places in south-eastern Australia (Moreton Island, Solitary Islands, Shellharbour, Eden) and New Zealand (Tawarahnui Peninsula, Mount Maunganui, Kaikoura and Milford Sound) (see Supplementary Information for place information). Samples were gathered by hand from rocky reefs by diving or snorkelling and either snap frozen in liquid nitrogen (as in Ref.10) or dried in silica gel (as in Ref.15). One specimen gathered at Mooloolaba, Queensland, 60 km north of the recognized extant series of E. radiata, was gathered as dried beach wreck. As the tasting techniques for herbarium specimens are mostly unidentified, we treat their geographical origin as that initially specified by the collector. All approaches were carried out in accordance with pertinent standards and guidelines; samples were gathered under clinical authorizations released by state or federal governments authorizations (P01/0059( A) -2.0 and QS2018/MAN471); and herbarium specimens were gotten under contracts with each herbarium/institution. DNA extraction and gene amplificationFor fresh, dried and protected herbarium specimens around 50 mg of tissue was utilized for extraction. For the single formalin repaired sample, 150 mg of internal stipe tissue was utilized for extraction as thallus tissue stopped working to produce DNA or magnify. DNA was separated utilizing (a) a combined CTAB and SDS protocol39 or (b) the Qiagen DNeasy Plant Pro package, following the maker’s guidelines. For both procedures the tissue lysis action was reached 10 days incubation at 65 ° C with samples vortexed two times daily. Drawn out DNA from fresh product was cleansed utilizing a Qiagen DNeasy PowerClean Pro clean-up package following the maker’s procedure. Cleansed DNA that was considered of adequate quality and amount (a minimum of ~ 3 ng/ µL with > 600 bp molecular weight) was PCR magnified for a part of the mitochondrial cytochrome c oxidase 1– 5 ′ barcoding area (COI) with the formerly released guides, Gaz F2 (5 ′ CCAACCAYAAAGATATWGGTAC 3 ′) and Gaz R2 (5 ′ GGATGACCAAARAACCAAAA 3 ′) 40 in 50 µL volumes utilizing the Invitrogen Platinum II Hot-start PCR Master-Mix (2x). To lower the threat of contamination, DNA extractions and PCRs were carried out in different locations with their own pipettes and devices, and PCR set-up was performed utilizing newly opened plans of consumables with all work performed in a UV-sterilised laminar circulation cabinet41,42. A PCR mixed drink was prepared based on the maker’s guidelines (7.5 µL design template) and cycled in an Eppendorf Mastercycler Nexus Gradient thermocycler. Visualisation of PCR items was performed utilizing a 2% agarose plate to figure out the response success for each marker. Numerous samples stopped working to magnify utilizing the Invitrogen Platinum II Hot-start package, so we likewise enhanced 1.0 µL of DNA extract in a repeat PCR in a Thermo Scientific Phire Plant Direct PCR Master Mix package following the maker’s ‘water down and save’ procedure. This package is developed to magnify DNA straight from plant tissues without the requirement for extraction and filtration. The polymerase is optimised to endure PCR inhibitors such as polyphenols and polysaccharides, and needs extremely percentages of DNA design template. We have actually discovered it to work well for poorer-quality, low-concentration, herbarium obtained DNA. In accordance with the Thermo Scientific Phire Plant Direct PCR Master Mix set maker directions, a gradient series PCR was carried out, utilizing cleansed DNA, to identify the ideal annealing temperature level for the primer/template set (53.5 ° C). PCR biking included a preliminary denaturation for 5 minutes at 98 ° C, followed by 45 cycles of: 5 s at 98 ° C, 5 s at 53.5 ° C and 45 s at 72 ° C. A last extension was performed for 1 minutes at 72 ° C. Successful amplicons that produced a clear band on a gel with a clear unfavorable control were contracted out for filtration and sequencing at the Australian Genomic Research Facility (AGRF), Sydney. We keep in mind that for the herbarium samples it typically took 2 to 3 repeat extractions and/or 3 to 4 PCR responses tweaking master blends, design template volumes and thermocycler conditions to get ideal item for sequencing. Forward and reverse series checks out were de novo put together utilizing Geneious Prime 11.1.541,42 and modified by eye. A more 76 COI series originated from E. radiata from the south-western Pacific26 were obtained from GenBank (Supplementary Information) and were lined up with the newly-generated series utilizing the MUSCLE algorithm consisted of in Geneious Prime utilizing default settings43. Forward and reverse guides were cut from series and 2 information quality checks were performed: a BLAST search44 to compare series with those kept in NCBI-accessible databases to validate that right hereditary markers had actually been magnified, and protein translation on the total positioning based upon the basic hereditary code. Haplotype variety and networkThe COI nucleotide positioning of Ecklonia radiata series was exported from Geneious Prime in NEXUS format for usage in haplotype variety analysis and restoration of an haplotype network. Variety was evaluated utilizing the DNA Sequence Polymorphism plan (DNAsp) 45 where samples were aggregated into local populations for visualisation and analysis based upon shared haplotype identity: South Australia, Victoria and Tasmania (SA, Vic Tas); New South Wales (NSW); Queensland (QLD); and Lord Howe Island, Norfolk Island, Kermadec Islands and New Zealand (LHI, NI, KI, NZ). A Templeton-Crandall-Sing (TCS) haplotype network46 was rebuilded utilizing PopART software47. The resultant network graphic was enhanced and overlaid on a vector map utilizing InkScape 1.148 Remote or deep-water areas made sample collection challenging, and for this factor sample numbers for some areas are low. As an outcome, conclusions based upon tasting from these places require to be related to with care since there might be unsampled haplotypes. Last glacial optimums dataTo show the most likely temperature-driven latitudinal shift in kelp distributional patterns in between the LGM and present, we utilized existing day quotes of kelp cover29 offered recognized links in between population size, hereditary variety, and the a priori presumption that areas with high kelp cover would likely have actually supported high hereditary diversity38,49,50. Here, the relationship in between kelp cover and indicate yearly sea surface area temperature level throughout eastern Australia (18– 38 ° S) was originated from field studies carried out over a ~ 1000 km latitudinal gradient and remotely-sensed ocean surface area temperature level (see Davis et al. 29). This relationship was utilized to forecast and compare the circulations of mean kelp cover approximated for the contemporary and the LGM. Contemporary cover was approximated utilizing a 25-year average of mean yearly sea surface area temperature level incorporating the duration 1996– 2020 sourced from the Copernicus Marine Environment Monitoring Service (https://marine.copernicus.eu; item # 010_011). Kelp cover at the LGM was approximated utilizing mean yearly sea surface area temperature level information rebuilded for the oceans around Australia utilizing planktonic foraminifera assemblages by Barrows and Juggins30. Approximated kelp cover for the contemporary and LGM were then balanced latitudinally within the optimum variety of depths (i.e. 0– 80 m2,14) that E. radiata is understood to take place. Considered that water level was roughly 120 m lower at the LGM, the analysis for this duration incorporated continental rack that is 120– 200 m deep at the contemporary water level (representing 0– 80 m depth at the LGM). This analysis does not take into account any cross-shelf irregularity in environment accessibility (rocky reef) that might have happened as sea level fell, due to unpredictability relating to regional geomorphological procedures and the subsequent circulation of rocky substrate throughout the LGM. We likewise acknowledge that aspects aside from ocean temperature level (e.g. nutrient and light schedule and biological interactions) add to the understood circulation of kelp cover29 and these were not integrated here. ResultsA overall of 43 COI series (658 bp) were newly-generated for this research study, representing the very first COI barcode dataset for E. radiata from northern NSW, southern Queensland, Lord Howe Island, Norfolk Island and New Zealand (Supplementary Information). MegaBLAST searches of newly-generated COI series (consisting of the historical herbarium samples) validated that all samples were E. radiata (> 98%). Variety and haplotype networksA overall of 6 haplotypes existed amongst E. radiata samples from the research study location (H1 and H8 to12) (Note: we follow Coleman et al. 6 in the allotment of haplotype numbers to keep clearness amongst Ecklonia radiata haplotypes, where H1 is extensively dispersed throughout the Southern Hemisphere and H8– 12, chosen here, are personal to the south-west Pacific. H 2– 7 are E. radiata haplotypes that are not present here however in Southern Africa and Oman6). Nucleotide distinctions amongst haplotypes were little (Table 1), the most varied population was in southern Queensland (Hd= 0.636) followed by Victoria, Tasmania and South Australia (Hd= 0.486) and NSW (Hd= 0.077). A single, extensively dispersed haplotype H1 (n= 74), was discovered throughout the whole research study location from south-eastern South Australia, south to western Tasmania, north along the NSW coast into southern QLD east to Norfolk Island (NI), the Kermadec Islands (KI), south to the north island of New Zealand (NZ) and at Lord Howe Island (LHI) (Fig. 1). Table 1 Diversity metrics for Ecklonia radiata in the south-western Pacific.Full size table One haplotype was limited to cool-temperate waters: H12 (n= 35) discovered in Bass Strait and south-eastern Tasmania (identified as H1 haplotype in Durrant et al. 26). On the other hand, 4 haplotypes were limited to subtropical waters north of 30 ° S. One haplotype (H8 (n= 1)) was discovered in northern NSW (Solitary Islands), 2 were from Moreton Island, QLD (H10 (n= 7) and H11 (n= 1)) and a single haplotype that was 3 base-pair replacements various from H1, H9 (n= 1), was discovered at Mooloolaba, QLD (Fig. 1). Forecasting cover of Ecklonia at the LGMModelling exposed that throughout the last glacial optimum the ideal mean yearly temperature level isotherm for E. radiata [~ 23 °C]29 might have lain in between Rockhampton (| 21 ° S) and Moreton Island (| 27 ° S) 30,51 and, constant with modern biomass distributions29, this area would likely have actually supported thick populations of E. radiata (Fig. 2). High typical kelp cover would have extended in between 31 and 24 ° S and have actually resembled extant areas around 34 ° S, a, equatorward shift of 3 to 10 degrees of latitude. Figure 2Estimated suggest cover (%) of Ecklonia radiata anticipated for the (a) last glacial optimum (LGM) based upon rebuilt mean yearly sea surface area temperature level information from Barrows and Juggins30, and (b) contemporary based upon a 25-year average of mean yearly sea surface area temperature level incorporating the duration 1996– 2020. Panel (c) shows the latitudinal average of approximated mean E. radiata cover at the LGM (blue information) and contemporary (black information) within the optimum variety of depths (0– 80 m) that kelp takes place. Spatial forecasts of mean E. radiata cover provided in panels (a, b) extend throughout the complete continental rack (i.e. 0– 200 m depth variety) to assist visual analysis of the latitudinal patterns provided, however information displayed in panel (c) show the approximated mean kelp cover within the optimum series of depths (0– 80 m) that kelp takes place. Relative to contemporary water level, the contemporary analysis (black information) incorporates the 0– 80 m part of the continental rack, while the LGM analysis (blue information) includes the 120– 200 m part of the continental rack (i.e. 0– 80 m at LGM water level). Complete size image DiscussionWarm, rear edge populations of temperate types can typically harbour high and special evolutionary hereditary variety as they represent historical refugia from previous climates11,20,21. We took a look at whether such deep-water rear-edge kelp populations off eastern Australia might have been refugia throughout durations of glaciation when water level fell, temperature level cooled and the accessibility of rocky reef decreased along this coast23,24,25. We discovered really high and distinct evolutionary variety in these deep, rear-edge populations of E. radiata compared to anywhere else tested throughout the circulation of this species6. This variety lived mostly in a little, separated population that is limited to deep water (> 30 m) off Moreton Island supporting the most likely function of this reef as a sanctuary under previous environments. Distinct variety was discovered in a single specimen of beach wreck 60 km north of the recognized extant variety of E. radiata in Mooloolaba, recommending that undiscovered deep-water populations might exist along this coast. This is supported by spatial modelling based upon ecophysiological and oceanographic information that showed that throughout the LGM, kelp forests were most likely to have actually been comprehensive and thick in south-eastern Queensland. Historic haven and circulation of Ecklonia radiata The reasonably high and distinct kelp hereditary variety off south-eastern Queensland (Moreton Island and Mooloolaba) recommends that these rocky reefs were likely a historical haven for temperate marine organisms under previous environments when waters were cooler and water level lower. Throughout the last glacial optimum (LGM; 21,000 ± 2000 cal year BP) the ideal mean yearly temperature level isotherm for E. radiata [~ 23 °C]29 was approximated to be situated in between Rockhampton (| 21 ° S) and Moreton Island (| 27 ° S) 30,52 and, constant with modern biomass distributions29, this area would likely have actually supported big and thick populations of E. radiata (Fig. 2). At this time, water level were roughly 65 to 125 m lower than present mean sea level25 and within 60 km of today coastline23 restricting E. radiata to the couple of rocky reefs that exist on the rack (e.g. China Wall, Henderson’s Reef) and locations with unconsolidated rocky substrates31. Even more, the majority of the rocky reef south of Moreton Island where the continental rack is really narrow (only|25 km broad) 53, would have emersed, restricting environment accessibility and limiting the circulations of rocky reef marine organisms along much of the NSW coast. The supremacy of haplotype H1 throughout much of the contemporary variety of E. radiata recommends quick recolonisation from refugia following glacial termination or variety contraction. Post-glacial warming would likely have actually allowed quick poleward dispersal and growth consisting of throughout the Tasman Sea past Lord Howe and Norfolk Islands to New Zealand into the contemporary latitudinal variety for E. radiata (some 10 ° of latitude south of LGM isotherms). The concomitant increase in water level of around 60– 120 m flooded big locations of what is now shallow continental rack waters off NSW where the recently readily available rocky benthos supplied area for the main succession of this leader genotype. As gone over by Hewitt54 leader genotypes (H1 haplotype) would control this brand-new variety and any subsequent genotypic dispersal is less most likely to develop due to optimum bring capability and competitors (i.e. high-density stopping) within the recognized populations52,55. As glaciers even more pulled away, and marine isotherms moved even more south helping with present day types circulations, the refugial E. radiata populations in southern Queensland would have ended up being progressively thermally stressed out over timescales of weeks to months as temperature levels went beyond the thermal limit for E. radiata (| 26 ° C2) other than at depth. 20 ° C )however quickly catch warmer temperature levels in spring and summer season when temperature levels go beyond|27 ° C( pers. obs. T. Stevens). Lord Howe ( 31 ° S, SST 19– 26 ° C), where E. radiata is putatively extinct, sits within the circulation of the warm east Australian present where temperature levels now primarily go beyond thermal tolerances in summer56. Norfolk Island (| 29 ° S) is somewhat cooler (SST 18– 25 ° C) which permits E. radiata to exist in separated deep-water environments. Studies for E. radiata in deep water refugia are necessitated on Lord Howe Island (to examine presence/absence) and Norfolk Island (to map geographical level), in order to even more illuminate how previous and present environment forms the circulation of this crucial kelp and to prioritise locations for preservation. On a wider scale, Ecklonia radiata populations in the Indian Ocean display comparable haplotype variety (Hd= 0.586, 7 haplotypes) to the south-Western Pacific (Hd= 0.526, 6 haplotypes). The H1 haplotype extends westward into the Indian Ocean from southern Western Australia, west to southern Africa, and, traditionally at Oman in the north6. In a number of variety edge populations at the Cape Province of South Africa and at the Abrolhos Islands in Western Australia, the extensive H1 is missing, changed by regionally-unique, personal haplotypes6 recommending these locations have actually been separated traditionally or physically (Abrolhos, Western Australia). These patterns in evolutionary variety contrast with modern patterns in neutral variety in this species15,38 and other macroalgal taxa9,57 which decrease with increasing latitude along the very same shoreline. This is because of the strong poleward circulation of the East Australian Current that isolates temperate macroalgal populations at low latitudes. Low variety at low latitudes can likewise be adaptive9,10 and these populations have loci under choice connected to greater and more variable temperature levels, consisting of those off Moreton Island (Minne et al. unpbl. information). Comprehending the relationships in between evolutionary and modern patterns of variety would gain from genotyping samples utilizing both COI and SNP markers in addition to thermal tension experiments on chosen haplotypes. While making use of herbarium collections to take a look at patterns of hereditary variety is starting to acquire traction for terrestrial plants58), this research study is among the couple of to effectively enhance and series DNA from old algal herbarium samples. Coleman et al. 6 effectively enhanced DNA from 47 years of age samples from Oman kept in 3 various herbaria. Goff and Moon59 enhanced DNA from 29 year old samples and Nahor et al. 60 from a 117 year old sample. We likewise effectively drawn out DNA from a formalin maintained sample. We keep in mind that this stays a difficult job. Effective DNA extraction and PCR amplification typically needed several efforts, tweaking both extraction and PCR procedures. We advise the Themo Fisher Phire Plant Direct PCR package which is created to enhance percentages of DNA in the existence of plant-derived PCR inhibitors. Drawing out DNA from formalin maintained specimens might be simpler utilizing the internal parts of the thicker thallus or stipe in which formalin does not permeate as deeply. Herbariums are an underutilised resource for analyzing historic genomic patterns yet hold terrific guarantee for revealing the motorists of seaweed circulation and environment impacts61. Contemporary environment modification is threatening the high and distinct hereditary variety discovered amongst eastern Australian low-latitude range-edge populations with warming triggering decreases of E. radiata along this coastline7. Forecasts recommend additional decreases and variety contractions of kelp under future environment scenarios29 as has actually taken place in variety edge E. radiata populations globally6,8. While it is not likely that in situ security might stop decreases of rear edge kelp populations under circumstances of warming, their special hereditary variety might be safeguarded and studied ex situ in culture banks for usage in remediation, hybridisation62 or helped adjustment methods. Data availabilityThe datasets created and/or evaluated throughout the present research study are readily available in the NCBI GenBank repository at https://www.ncbi.nlm.nih.gov/nuccore. GenBank accession numbers for each series are offered in Supplementary Information. ReferencesKrumhansl, K. A. et al. International patterns of kelp forest modification over the previous half-century. Proc. Natl. Acad. Sci. 113( 48 ), 13785– 13790. https://doi.org/10.1073/pnas.1606102113 (2016 ). Post ADS CAS Google Scholar Wernberg, T. et al. Biology and ecology of the internationally substantial kelp Ecklonia radiata. Oceanogr. Mar. Biol. https://doi.org/10.1201/9780429026379-6( 2019). Short Article Google Scholar Bennett, S. et al. The ‘Great Southern Reef’: Social, environmental and financial worth of Australia’s overlooked kelp forests. Mar. Freshw. Res. 67( 1), 47– 56. https://doi.org/10.1071/MF15232( 2015 ). Short Article Google Scholar Eger, A. et al. The financial worth of fisheries, blue carbon, and nutrient biking in international marine forests. EcoEvoRxiv. https://doi.org/10.32942/osf.io/n7kjs( 2021). Post Google Scholar Smith, K. E. et al. Socioeconomic effects of marine heatwaves: Global concerns and chances. Science 374, 6566. https://doi.org/10.1126/science.abj3593 (2021). Post CAS Google Scholar Coleman, M. et al. Loss of a internationally special kelp forest and hereditary variety from the northern hemisphere. Sci. Rep. 12, 5020. https://doi.org/10.1038/s41598-022-08264-3 (2022 ). Short Article ADS CAS Google Scholar Vergés, A. et al. Long-lasting empirical proof of ocean warming resulting in tropicalization of fish neighborhoods, increased herbivory, and loss of kelp. Proc. Natl. Acad. Sci. 113( 48 ), 13791– 13796. https://doi.org/10.1073/pnas.1610725113 (2016 ). Post ADS CAS Google Scholar Wernberg, T. et al. Climate-driven routine shift of a temperate marine environment. Science 353( 6295 ), 169– 172. https://doi.org/10.1126/science.aad8745( 2016). Post ADS CAS Google Scholar Wood, G. et al. Genomic vulnerability of a dominant seaweed indicate future-proofing paths for Australia’s undersea forests. Glob. Modification Biol. 27( 10 ), 2200– 2212. https://doi.org/10.1111/gcb.15534( 2021). Post ADS CAS Google Scholar Vranken, S. et al. Genotype-environment inequality of kelp forests under environment modification. Mol. Ecol. 30 (15 ), 3730. https://doi.org/10.1111/mec.15993( 2021). Post Google Scholar Assis, J. et al. Deep reefs are weather refugia for hereditary variety of marine forests. J. Biogeogr. 43( 4), 833– 844. https://doi.org/10.1111/jbi.12677( 2016). Post Google Scholar Lourenço, C. R. et al. Upwelling locations as environment modification refugia for the circulation and hereditary variety of a marine macroalga. J. Biogeogr. 43( 8), 1595– 1607. https://doi.org/10.1111/jbi.12744( 2016). Short Article Google Scholar Graham, M. H., Kinlan, B. P., Druehl, L. D., Garske, L. E. & Banks, S. Deep-water kelp refugia as possible hotspots of tropical marine variety and performance. Proc. Natl. Acad. Sci. 104 (42), 16576– 16580. https://doi.org/10.1073/pnas.0704778104 (2007). Post ADS Google Scholar Marzinelli, E. M. et al. Massive geographical variation in circulation and abundance of Australian deep-water kelp forests. PLoS ONE 10, e0118390. https://doi.org/10.1371/journal.pone.0118390 (2015 ). Post CAS Google Scholar Coleman, M. A. et al. Variation in the strength of continental border currents figures out continent-wide connection in kelp. J. Ecol. 99( 4), 1026– 1032. https://doi.org/10.1111/j.1365-2745.2011.01822.x (2011). Post Google Scholar Hampe, A. & Petit, R. J. Conserving biodiversity under environment modification: The rear edge matters. Ecol. Lett. 8( 5), 461– 467. https://doi.org/10.1111/j.1461-0248.2005.00739.x (2005 ). Short article Google Scholar Maggs, C. A. et al. Examining signatures of glacial refugia for North Atlantic benthic marine taxa. Ecology 89( sp11 ), S108– S122. https://doi.org/10.1890/08-0257.1 (2008 ). Post Google Scholar Grant, W. S., Lydon, A. & Bringloe, T. T. Phylogeography of split kelp Hedophyllum nigripes: Northern ice-age refugia and trans-Arctic dispersal. Polar Biol. 43, 1829– 1841. https://doi.org/10.1007/s00300-020-02748-6( 2020). Post Google Scholar Hoarau, G., Coyer, J. A., Veldsink, J. H., Stam, W. T. & Olsen, J. L. Glacial refugia and recolonization paths in the brown seaweed Fucus serratus. Mol. Ecol. 16( 17), 3606– 3616. https://doi.org/10.1111/j.1365-294X.2007.03408.x (2007). Post CAS Google Scholar Fraser, C. I., Nikula, R., Spencer, H. G. & Waters, J. M. Kelp genes expose results of subantarctic sea ice throughout the Last Glacial Maximum. Proc. Natl. Acad. Sci. 106 (9 ), 3249– 3253. https://doi.org/10.1073/pnas.0810635106( 2009 ). Short Article ADS Google Scholar Assis, J. et al. Previous environment modifications and strong oceanographic barriers structured low-latitude hereditary antiques for the golden kelp Laminaria ochroleuca. J. Biogeogr. 45( 10), 2326– 2336. https://doi.org/10.1111/jbi.13425( 2018). Post Google Scholar Gersonde, R., Crosta, X., Abelmann, A. & Armand, L. Sea-surface temperature level and sea ice circulation of the Southern Ocean at the EPILOG last glacial optimum– A circum-Antarctic view based upon siliceous microfossil records. Quat. Sci. Rev. 24( 7– 9), 869– 896. https://doi.org/10.1016/j.quascirev.2004.07.015( 2005 ). Short Article ADS Google Scholar Bostock, H. C., Opdyke, B. N., Gagan, M. K., Kiss, A. E. & Fifield, L. K. Glacial/interglacial modifications in the East Australian existing. Clim. Dyn. 26, 645– 659. https://doi.org/10.1007/s00382-005-0103-7( 2006). Short Article Google Scholar Brooke, B. P., Nichol, S. L., Huang, Z. & Beaman, R. J. Palaeoshorelines on the Australian continental rack: Morphology, sea-level relationship and applications to ecological management and archaeology. Cont. Rack Res. 134, 26– 38. https://doi.org/10.1016/j.csr.2016.12.012 (2017). Short Article ADS Google Scholar Williams, A. N., Ulm, S., Sapienza, T., Lewis, S. & Turney, C. S. M. Sea-level modification and demography throughout the last glacial termination and early Holocene throughout the Australian continent. Quat. Sci. Rev. 182, 144– 154. https://doi.org/10.1016/j.quascirev.2017.11.030( 2018 ). Short Article ADS Google Scholar Durrant, H. M. S., Barrett, N. S., Edgar, G. J., Coleman, M. A. & Burridge, C. P. Shallow phylogeographic histories of crucial types in a biodiversity hotspot. Phycologia 54( 6), 556– 565. https://doi.org/10.2216/15-24.1 (2015 ). Short Article Google Scholar O’Hara, T. D. & Poore, G. C. B. Patterns of circulation for southern Australian marine echinoderms and decapods. J. Biogeogr. 27( 6), 1321– 1335. https://doi.org/10.1046/j.1365-2699.2000.00499.x (2000 ). Short article Google Scholar Waters, J. M. Marine biogeographical disjunction in temperate Australia: Historical landbridge, modern currents, or both? Scuba divers. Distrib. 14( 4 ), 692– 700. https://doi.org/10.1111/j.1472-4642.2008.00481.x (2008 ). Short article Google Scholar Davis, T. R., Champion, C. & Coleman, M. A. Climate refugia for kelp within an ocean warming hotspot exposed by stacked types circulation modelling. Mar. Environ. Res. 166, 105267. https://doi.org/10.1016/j.marenvres.2021.105267(2021 ). Short article CAS Google Scholar Barrows, T. T. & Juggins, S. Sea-surface temperature levels around the Australian margin and Indian Ocean throughout the last glacial optimum. Quat. Sci. Rev. 24(7– 9), 1017– 1047. https://doi.org/10.1016/j.quascirev.2004.07.020 (2005 ). Post ADS Google Scholar Richmond, S. & Stevens, T. Classifying benthic biotopes on sub-tropical continental rack reefs: How beneficial are abiotic surrogates? Estuar. Coast. Rack Sci. 138, 79– 89. https://doi.org/10.1016/j.ecss.2013.12.012 (2014 ). Post ADS Google Scholar Jordan, A. et al. Seabed Habitat Mapping of the Continental Shelf of NSW (New South Wales Department of Environment, Climate Change and Water, 2010). Google Scholar Lewis, S. E., Sloss, C. R., Murray-Wallace, C. V., Woodroffe, C. D. & Smithers, S. G. Post-glacial sea-level modifications around the Australian margin: An evaluation. Quat. Sci. Rev. 74, 115– 138. https://doi.org/10.1016/j.quascirev.2012.09.006 (2013 ). Post ADS Google Scholar Millar, A. J. K. Marine benthic algae of Norfolk island, South Pacific. Aust. Syst. Bot. 12( 4 ), 479– 547. https://doi.org/10.1071/SB98004 (1999 ). Short article Google Scholar Ridgway, K. R. & Dunn, J. R. Mesoscale structure of the mean East Australian existing system and its relationship with topography. Prog. Oceanogr. 56, 189– 222. https://doi.org/10.1016/S0079-6611( 03 )00004-1 (2003 ). Short article ADS Google Scholar Lough, J. M. & Hobday, A. J. Observed environment modification in Australian marine and freshwater environments. Mar. Freshw. Res. 62(9), 984– 999. https://doi.org/10.1071/MF10272(2011). Post Google Scholar Sunday, J. M. et al. Types characteristics and environment speed describe geographical variety shifts in an ocean-warming hotspot. Ecol. Lett. 18( 9 ), 944– 953. https://doi.org/10.1111/ele.12474 (2015 ). Post Google Scholar Coleman, M. A. et al. Variation in the strength of continental limit currents figures out patterns of massive connection in kelp. J. Ecol. 99, 1026– 1032 (2011 ). Short article Google Scholar Maeda, T., Kawai, T., Nakaoka, M. & Yotsukura, N. Effective DNA extraction technique for piece analysis utilizing capillary sequencer of the kelp, Saccharina. J. Appl. Phycol. 25, 337– 347. https://doi.org/10.1007/s10811-012-9868-3 (2013 ). Post CAS Google Scholar Lane, C. E., Lindstrom, S. C. & Saunders, G. W. A molecular evaluation of northeast Pacific Alaria types (Laminariales, Phaeophyceae) with referral to the energy of DNA barcoding. Mol. Phylogenet. Evol. 44( 2 ), 634– 648. https://doi.org/10.1016/j.ympev.2007.03.016 (2007 ). Short article CAS Google Scholar Saunders, G. W. & McDevit, D. C. Acquiring DNA series information from dried archival red algae (Florideophyceae) for the function of using offered names to modern hereditary types: An important evaluation. Botany 90, 191– 203 (2012 ). Post CAS Google Scholar Kearse, M. et al. Geneious standard: An incorporated and extendable desktop software application platform for the company and analysis of series information. Bioinformatics 28( 12 ), 1647– 1649. https://doi.org/10.1093/bioinformatics/bts199 (2012 ). Short article Google Scholar Edgar, R. C. MUSCLE: numerous series positioning with high precision and high throughput. Nucleic Acids Res. 32( 5 ), 1792– 1797. https://doi.org/10.1093/nar/gkh340 (2004 ). Post CAS Google Scholar Altschul, S. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. J. Basic regional positioning search tool. J. Mol. Biol. 215( 3 ), 403– 410. https://doi.org/10.1016/S0022-2836( 05 )80360-2 (1990 ). Post CAS Google Scholar Rozas, J. et al. DnaSP 6: DNA series polymorphism analysis of big information sets. Mol. Biol. Evol. 34( 12 ), 3299– 3302. https://doi.org/10.1093/molbev/msx248 (2017 ). Post CAS Google Scholar Clement, M., Posada, D. & Crandall, K. A. TCS: A computer system program to approximate gene genealogies. Mol. Ecol. 9( 10 ), 1657– 1659. https://doi.org/10.1046/j.1365-294x.2000.01020.x (2000 ). Post CAS Google Scholar Leigh, J. & Bryant, D. PopART: Full-feature software application for haplotype network building and construction. Approaches Ecol. Evol. 6( 9 ), 1110– 1116. https://doi.org/10.1111/2041-210X.12410 (2015 ). Short article Google Scholar Inkscape Project. Inkscape Project. https://inkscape.org/ (2020 ). Coleman, M. A. et al. Connection within and amongst a network of temperate marine reserves. PLoS ONE 6( 5 ), e20168. https://doi.org/10.1371/journal.pone.0020168 (2011 ). Post ADS CAS Google Scholar Davis, T. R., Cadiou, G., Champion, C. & Coleman, M. A. Environmental chauffeurs and signs of modification in environment and fish assemblages within an environment modification hotspot. Reg. Mar. Stud. https://doi.org/10.1016/j.rsma.2020.101295(2020). Short Article Google Scholar Mix, A. C., Bard, E. & Schneider, R. Environmental procedures of the glacial epoch: Land, oceans, glaciers( EPILOG). Quat. Sci. Rev. 20 (4), 627– 657. https://doi.org/10.1016/S0277-3791(00)00145-1( 2001 ). Short article ADS Google Scholar Waters, J. M. Competitive exemption: Phylogeography’s ‘elephant in the space’? Mol. Ecol. 20( 21 ), 4388– 4394. https://doi.org/10.1111/j.1365-294X.2011.05286.x (2011 ). Short article Google Scholar Cresswell, G. R., Peterson, J. L. & Pender, L. F. The East Australian present, upwellings and downwellings off eastern-most Australia in summer season. Mar. Freshw. Res. 68 (7 ), 1208– 1223. https://doi.org/10.1071/MF16051 (2016 ). Short article Google Scholar Hewitt, G. Some hereditary repercussions of glacial epoch, and their function in divergence and speciation. Biol. J. Linn. Soc. 58( 3 ), 247– 276. https://doi.org/10.1006/bijl.1996.0035 (1995 ). Post Google Scholar Waters, J. M., Fraser, C. I. & Hewitt, G. M. Founder takes all: Density-dependent procedures structure biodiversity. Patterns Ecol. Evol. 28( 2 ), 78– 85. https://doi.org/10.1016/j.tree.2012.08.024 (2013 ). Post Google Scholar Wernberg, T. et al. Hereditary variety and kelp forest vulnerability to weather tension. Sci. Rep. 8( 1851 ), 1– 8. https://doi.org/10.1038/s41598-018-20009-9 (2018 ). Short article CAS Google Scholar Coleman, M. A. & Kelaher, B. P. Connectivity amongst fragmented populations of a habitat-forming alga, Phyllospora comosa (Phaeophyceae, Fucales) on an urbanised coast. Mar. Ecol. Prog. Ser. 381, 63– 70 (2009 ). Post ADS Google Scholar Drábková, L. Z. DNA extraction from herbarium specimens. In Molecular Plant Taxonomy. Approaches in Molecular Biology Vol. 1115 (ed. Besse, P.) (Humana Press, 2014). Google Scholar Goff, L. J. & Moon, D. A. PCR amplification of nuclear and plastid genes from algal herbarium specimens and algal spores 1. J. Phycol. 29, 381 (1993 ). Post CAS Google Scholar Nahor, O., Luzzatto-Knaan, T. & Israel, A. A brand-new hereditary family tree of Asparagopsis taxiformis (Rhodophyta) in the Mediterranean Sea: As the DNA barcoding shows a current Lessepsian intro. Front. Mar. Sci. https://doi.org/10.3389/fmars.2022.873817(2022). Post Google Scholar Coleman, M. A. & Brawley, S. H. Variability in temperature level and historic patterns in recreation in the Fucus distichus complex (Heterokontophyta; Phaeophyceae): Implications for speciation and collection of herbarium specimens. J. Phycol. 41, 1110– 1119(2005). Post Google Scholar Martins, N. et al. Hybrid vigour for thermal tolerance in hybrids in between the allopatric kelps Laminaria digitata and L. pallida(Laminariales, Phaeophyceae) with contrasting thermal affinities. Eur. J. Phys. 54 (4), 548– 561(2019). CAS Google Scholar Download referrals AcknowledgementsThe authors thank 4 customers for remarks which considerably enhanced the manuscript. They likewise thank R. Lawton from University of Waikato, S. Hart and P. Hart and the Auckland War Memorial Museum( Tāmaki Paenga Hira), for supplying Ecklonia radiata from New Zealand; the personnel at the National Herbarium of Melbourne, New South Wales and Queensland Herbarium for supplying tissue samples from Australian specimens. MAC and TW were supported by the Australian Research Council (LP190100346, DP200100201). Author informationAuthors and AffiliationsNew South Wales Department of Primary Industries, National Marine Science Centre, Coffs Harbour, NSW, Australia Matt J. Nimbs, Tom R. Davis, Curtis Champion & Melinda A. Coleman National Marine Science Centre, Southern Cross University, Coffs Harbour, NSW, Australia Matt J. Nimbs, Tom R. Davis, Curtis Champion & Melinda A. Coleman Oceans Institute and School of Biological Sciences, University of Western Australia, 35 Stirling Highway, Crawley, WA, 6009, Australia Thomas Wernberg & Melinda A. Coleman ContributionsM.A.C. and T.W. developed the concept, got main financing and carried out the fieldwork. M.A.C., T.M. and M.J.N. contributed samples and carried out parts of the lab work. C.C. and T.R.D. performed ecological condition analysis and produced figures. M.J.N. and M.A.C. carried out the analyses. M.J.N. and M.A.C. composed the manuscript. All authors added to moneying parts of the work and modifying the manuscript. Corresponding authorCorrespondence to Matt J. Nimbs. Principles statements Competing interests The authors state no completing interests. Extra informationPublisher’s noteSpringer Nature stays neutral with regard to jurisdictional claims in released maps and institutional associations. Additional InformationRights and approvals Open Access This post is certified under a Creative Commons Attribution 4.0 International License, which allows usage, sharing, adjustment, circulation and recreation in any medium or format, as long as you provide suitable credit to the initial author(s)and the source, supply a link to the Creative Commons licence, and show if modifications were made. The images or other 3rd party product in this post are consisted of in the post’s Creative Commons licence, unless suggested otherwise in a credit limit to the product. If product is not consisted of in the post’s Creative Commons licence and your planned usage is not allowed by statutory guideline or surpasses the allowed usage, you will require to get authorization straight from the copyright holder. To see a copy of this licence, see http://creativecommons.org/licenses/by/4.0/. Reprints and Permissions About this short article Cite this articleNimbs, M.J., Wernberg, T., Davis, T.R. et al. Environment modification threatens distinct evolutionary variety in Australian kelp refugia. Sci Rep 13, 1248(2023). https://doi.org/10.1038/s41598-023-28301-z Download citation Received: 28 March 2022 Accepted: 17 January 2023 Published: 23 January 2023 DOI: https://doi.org/10.1038/s41598-023-28301-z
- Thu. Jun 25th, 2026

Latest News